

1. Préambule
Cette synthèse s'articulera en deux parties : la première traitant de la calcification à l'échelle du polype, des cellules et des molécules, la deuxième partie traitera des données de calcification, de l'échelle de la colonie jusqu’à celle du récif. Cet article est une version francisée et vulgarisée d'un chapitre (Coral calcification, cells to reefs) du livre (Coral reef : an ecosystem in transition Dubinsky, Z et N. Stambler (Eds.), 2011 ) écrit par des chercheurs du Centre Scientifique de Monaco : Denis Allemand, Eric Tambutté, Didier Zoccola et Sylvie Tambutté, leaders mondiaux incontestés dans l'étude de la calcification des coraux.
2. Introduction
L'étude de la calcification chez les coraux scléractiniaires a débuté il y a maintenant plus d'un siècle et demi grâce aux travaux de Dana (1846). Dans un premier temps à but taxonomique, il a fallu attendre le milieu du XXème siècle pour que la physiologie de la calcification soit réellement abordée. Malgré plus de 150 ans d'études, force est de constater que les inconnues restent nombreuses comme l'attestent les deux citations ci-dessous, bien qu'elles soient issues de textes publiés avec 100 ans d'écart. "La question ayant un intérêt majeur à la fois pour les zoologistes et les paléontologistes, concerne les relations existant entre la partie molle du polype et la partie calcaire ou cornée secrétée par la plupart des coraux (Ogilvie 1887)". "La faible compréhension des mécanismes de calcification chez les coraux résulte du manque d'informations sur les interactions tissu/squelette et sur les patterns spatiaux et temporels de la morphogénèse" (Le Tissier 1987).
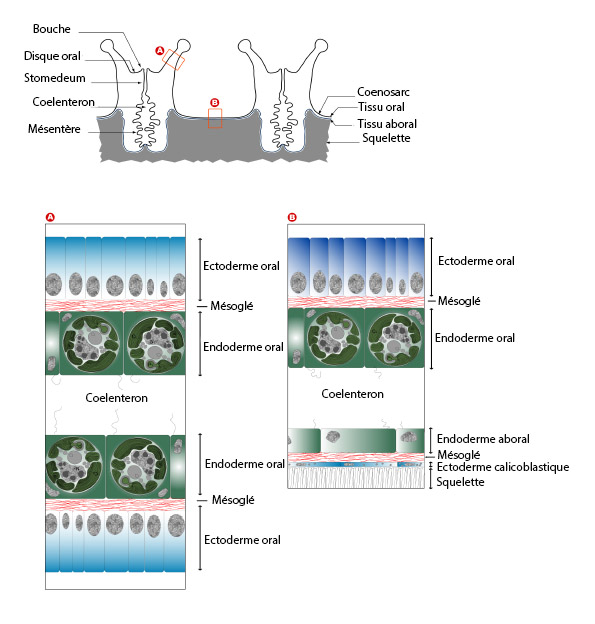
La partie vivante du corail est constituée par les polypes et entre eux le cœnosarque. Les coraux sont des organismes diploblastiques, donc organisés en deux feuillets cellulaires, l'ectoderme et l'endoderme, séparés par une mince couche de gelée (mésoglée). Lorsque les feuillets cellulaires font face à l'eau de mer on parle de tissus oraux (ectoderme oral, mésoglée orale et endoderme oral). Lorsque ces feuillets font face au squelette on parle de tissus aboraux (ectoderme aboral ou calicoblastique, mésoglée aborale et endoderme aboral).
3. Les différents types de biominéralisation et la calcification corallienne
Deux types de biominéralisation ont été décrits. Tout d'abord la minéralisation biologiquement induite, qui correspond aux processii dans lesquels aucune cellule spécialisée ou machinerie moléculaire (protéines) n'est utilisée spécifiquement pour la minéralisation. Les cristaux formés sont alors similaires à ceux obtenus de façon inorganique (un précipité par exemple). C'est un processus classique chez de nombreuses algues ou champignons par exemple. Le deuxième type de biominéralisation est dit "biominéralisation conduite par une matrice organique", renommé par la suite comme étant la "minéralisation biologiquement contrôlée".
Bien qu'encore légèrement discuté, la plupart des scientifiques sont d'accord pour admettre que le squelette corallien est issu d'un mécanisme biologiquement contrôlé. Ce contrôle trouvant naissance à deux niveaux différents au sein de l'organisme. Au niveau de la matrice organique, et/ou au niveau du transport des ions et de leur livraison au site de calcification. Ces deux niveaux de contrôle vont maintenant être abordés plus en détails.
3. Le site de la calcification corallienne : le milieu extracellulaire subcalicoblastique
L'anatomie des tissus coralliens est généralement comparée à un sac (le polype), attaché au squelette par sa base (Figure 1). Les tissus oraux font face à l'eau de mer, alors que les tissus aboraux font face au squelette. Chaque tissu est composé de deux feuillets cellulaires (monocouche cellulaire), l'ectoderme, et l'endoderme, respectivement pour la partie externe et interne du tissu. Ces deux feuillets sont séparés l'un de l'autre par une substance acellulaire et gélatineuse, la mésoglée. La cavité délimitée par les endodermes est le cœlenteron. Il correspond à la cavité gastrique.
3.1. Les tissus squeletogéniques : l'épithélium calicoblastique
Le tissu responsable de la calcification est l'épithelium calicoblastique, autrement appelé tissu squeletogène, ou calicodermique. Ce tissu est mis en place au moment de la métamorphose (passage de la vie pélagique larvaire à celle de vie fixée). Il est composé de deux types de cellules, les calicoblastes et les desmocytes. Ces derniers, de forme triangulaire sont impliqués dans l'ancrage des tissus au squelette, alors que les calicoblastes sont responsables de la calcification. La forme des calicoblastes est assez variable. Pouvant être fins et allongés, ils peuvent également former des sortes de poches selon l'activité de calcification. En effet, une forme plane est généralement associée à une calcification de faible activité, alors que les formes « en sac » sont généralement signes d'une forte activité de calcification. Dans ce dernier cas, il existe une correspondance stricte entre la forme de la cellule et de la fibre squelettique. Bien qu'une fibre soit en réalité formée par deux cellules calicoblastiques, cette forte correspondance peut être imagée par celle d'un moule et de la moulure qui en ressort.
3.2. Le milieu extracellulaire subcalicoblastique
De façon générale, un espace de largeur assez variable réside entre le tissu calicoblastique et le squelette, c'est l'espace subcalicoblastique. Cet espace peut présenter une épaisseur variable selon les espèces, ou encore l'heure de la journée. Il a été rapidement proposé que la calcification prenait place au sein de poches créées par un élargissement de l'espace subcalicoblastique. Et bien que l'épaisseur de cet espace au moment de la calcification soit encore largement étudiée, il semble bien admis que la calcification prend naissance dans cet espace au sein d'une matrice (milieu) gélatineuse secrétée par le tissu calicoblastique. Ce milieu calcifiant extracellulaire est probablement composé de protéines hydratées et de mucopolysaccharide (enchainement de sucres reliés les uns aux autres) qui forment une sorte d'hydrogel, où aurait lieu la première étape de la calcification : La nucléation.
3.3. Le milieu calcifiant extracellulaire, un compartiment ouvert ou fermé ?
Cette question peut sembler de prime abord relativement peu importante, cependant elle prend une toute autre facette lorsque l'on sait que les cellules d'un tissu peuvent être liées les unes aux autres par deux types de jonctions. Les jonctions serrées, ou jonctions septées dans le cas des invertébrés, permettent de former des barrières épithéliales totalement étanches. Dans ce cas, le milieu calcifiant extracellulaire serait un compartiment fermé. Le transport du matériel nécessaire à la calcification, le Ca2+ et le CO32- devrait donc se faire obligatoirement de façon transcellulaire, c'est à dire au travers de la cellule via des protéines spécialisées de type pompe ou canal. L'autre type de jonction est la jonction communicante qui permet de laisser passer de façon sélective certaines molécules en fonction de leur taille ou de leur charge. Ici, le milieu calcifiant extracellulaire serait ouvert, et le matériel nécessaire à la calcification pourrait passer entre les cellules. Dans le cas des coraux, seules les jonctions septées ont pu être physiquement misent en évidence. Cependant, diverses expérimentations ont montré que le tissu calicoblastique présentait une certaine perméabilité à différentes molécules. Il est donc impossible, au jour d'aujourd'hui de conclure sur ce point, mais il est tout à fait envisageable que le milieu calcifiant extracellulaire soit à la fois fermé et ouvert, selon le type de molécules candidates à la traversée de la barrière épithéliale.
3.4. Les caractéristiques physico-chimiques du milieu calcifiant extracellulaire
Déterminer la concentration ionique et le pH au site de calcification, apparait indispensable pour comprendre finement les mécanismes de biominéralisation. Cependant il n'existe à ce jour qu'une seule publication donnant partiellement de telles informations. Ainsi, le milieu calcifiant extracellulaire (MCE) a un pH plus basique (9,2 le jour et 8,1 la nuit) que celui mesuré au niveau des tissus (8,6 à 7,3) de jour comme de nuit, et une concentration en Ca2+ plus élevée de 0,58 mmol/L (jour) à 0,21 mmol/L (nuit) par comparaison à l'eau de mer. Il est important de noter que de tels écarts de concentration ne peuvent être permis sans le concours de mécanismes de transport actifs, et donc de consommateurs d'énergie. Il est également important de prendre ces données avec recul, car les seules méthodes utilisées jusqu'à aujourd'hui pour effectuer de telles mesures sont invasives, et nécessitent la perforation des tissus afin d'y insérer des microélectrodes. Cependant, des données issues d'approches indirectes semblent confirmer ces résultats, et attestent notamment d'une sursaturation de l'aragonite dans le MCE au site de calcification. Cette sursaturation a par ailleurs été estimée à un minimum de 7 fois supérieur à la valeur prise par l'eau de mer. Même en faible nombre, ces données suggèrent que la composition du MCE est contrôlée par le corail, et qu'il est donc isolé du milieu extérieur grâce à une perméabilité faible de l'ectoderme calicoblastique. Les données concernant les concentrations en CO2 (dioxyde de carbone) / HCO3- (hydroénocarbonate) / CO32- (carbonate) sont malheureusement inexistantes. Cependant, si l'on se réfère à ce qui est connu dans d'autres organismes biominéralisateurs (mollusque, oursin, utérus d'oiseaux...) il semble évident qu'ici aussi le carbone inorganique soit présent à des concentrations supérieures, comparé aux autres compartiments biologiques de l'organisme.
4. Physiologie de la calcification corallienne
Le squelette corallien est composé de carbonate de calcium, qui est précipité sous forme de cristaux d'aragonite. Comme nous l'avons vu précédemment, cette précipitation biocontrolée est localisée au niveau du MCE. Calcium et carbonate doivent donc y être apportés. Se pose alors la question de la provenance de ces éléments. Concernant le Ca2+, il provient naturellement du milieu extérieur, mais les choses ne sont pas si simples concernant le carbonate. En effet plusieurs origines sont possibles. Il peut, comme le Ca2+, provenir du milieu extérieur (sous forme de HCO3-, forme majoritairement présente au pH intracellulaire), mais aussi du CO2 métaboliquement produit par la respiration. La production de CO2 métabolique est d'ailleurs 9 fois supérieure à la quantité de carbone inorganique nécessaire à la calcification.
La provenance, mais surtout la forme chimique de ce carbone inorganique, implique des modifications notoires au niveau du bilan de la calcification.
Réaction de calcification
Ca2+ + CO32- => CaCO3
Réaction de calcification avec comme source de carbone inorganique du HCO3-
Ca2+ + HCO3- => CaCO3 + H+
Réaction de calcification avec comme source de carbone inorganique du CO2
Ca2+ + CO2 + H20 => CaCO3 + 2H+
Comme on peut le voir dans les équations chimiques mettant en jeu HCO3- et CO2 , des ions H+ sont des sous-produits de la réaction. Or ces ions acides doivent obligatoirement être rejetés hors du MCE afin d'y maintenir des conditions de pH permettant la précipitation du CaCO3 . La différence entre l'élimination d'un H+ ou de deux H+ peut sembler minime, mais le cout pour la cellule en est très loin. À ce jour, il est impossible d'affirmer laquelle de ces sources de carbone inorganique est utilisée. Toutefois, de nombreux indices tendent à montrer que chez Stylophora pistillata HCO3- serait préféré.
5. L'apport d'ions et leur élimination pour la calcification : une voie inactive et paracellulaire, ou active et transcellulaire
Au jour d'aujourd'hui, 4 hypothèses expliquent les voies utilisées pour l'apport d'ions au site de calcification:
- La première propose que les ions soient apportés au MCE en passant entre les cellules (voie paracellulaire) de l'ectoderme calicoblastique de façon passive (sans utilisation d'énergie). Ils diffusent donc selon leur gradient de concentration (du compartiment le plus concentré au moins concentré, ou de leur charge positive ou négative).
- La deuxième hypothèse propose que de l'eau de mer concentrée soit apportée au site de calcification. De la même façon que de l'ultrafiltration, les contractions du polype induiraient une ultrafiltration de l'eau de mer, et laisseraient passer de façon sélective les molécules de Ca2+ et de HCO3- au site de calcification (voie paracellulaire passive). Dans le même temps, le Ca2+ serait amené à sursaturation dans le MCE par le concours d'une voie transcellulaire (au travers des cellules) qui, via l'utilisation de protéines de type canal, permettrait d'amener du Ca2+ contre son gradient de concentration. Cette voie est dite active, car nécessite de l'énergie pour que les protéines canal fonctionnent et sursaturent le Ca2+ dans le MCE. L'utilisation d'énergie étant liée à l'activité ATPase de ces protéines
- La troisième hypothèse propose que seule la voie transcellulaire soit utilisée. Les ions seraient absorbés par des protéines canal au niveau de la face de la cellule opposée au squelette, puis transportés au travers de la cellule, et enfin transférés dans le MCE, via le même type de protéines canal. Cette voie est dite active, et est consommatrice d'énergie.
- La quatrième hypothèse est un mélange des voies para et transcellulaires. Ainsi, Ca2+ et HCO3- seraient apportés au MCE via une diffusion de type transcellulaire, mais aussi via l'absorption de ces derniers et leur livraison au MCE, en passant au travers de la cellule grâce à des protéines canal.
Comme d'habitude il est encore impossible de trancher entre ces différentes hypothèses, même si de nombreux indices montrent la présence d'un transport actif, mais aussi passif, et donc jouent en faveur des hypothèses 2 et 4.

CCe : Cellule calicoblastique; MCE : Milieu Calcifiant extracellulaire; Sq : squelette; PMO : protéine de la matrice organique; JS : Jonction septé; Pointillé rouge matrice organique; Ovale vert: Pompe ou protéine canal transportant les HCO3-; Ovale bleu: pompe ou protéine canal transportant le Ca2+.
6. Coût énergétique du transport d'ion
Comme cela à déjà été présenté précédemment, les taux de calcification des coraux sont plus importants le jour que la nuit. A travers diverses expériences utilisant des inhibiteurs spécifiques de certaines voies métaboliques, certains auteurs ont mis en évidence un lien direct entre métabolisme énergétique et calcification. Cependant, ce lien n'est vraiment significatif que lors des périodes de forte calcification (jour). Il a été montré que même si la calcification nocturne est de moindre importance, celle-ci reste biologiquement contrôlée, et n'est en aucun cas le fruit d’une précipitation que l'on pourrait caractériser d'hasardeuse. Des expériences complémentaires ont permis de montrer que calcification diurne et nocturne étaient toutes deux consommatrices d'énergie provenant du métabolisme de l'hôte et/ou de son symbiote. Ces résultats tendent par ailleurs à confirmer le caractère actif du transport des ions destinés aux processii de calcification.
7. Le rôle clef de la matrice organique
Le squelette des coraux et de nombreux biominéralisateurs ne contiennent pas que du CaCO3. En effet, il est également composé d'une partie organique appelée « matrice organique » (MO). On trouve cette matrice, à la fois entre et dans les cristaux, où elle sera protégée des agressions du temps durant plusieurs centaines d'années. La présence de cette MO est également un élément clef indiquant le contrôle biologique de la calcification. La composition exacte et le rôle de cette matrice sont encore débattus. Ainsi, dans les domaines biomédicaux où des fragments de squelettes coralliens sont utilisés en tant que bio-implant, de nombreuses publications la considèrent comme étant composée d’acides aminés libres. Pour les Géochimistes, elle ne serait que des restes de tissus emprisonnés lors de la précipitation.
7.1. Taux de MO dans le squelette
Chez les coraux, l'identification d'une MO date du 19eme siècle, où il était estimé qu'elle composait 4 à 8 % du squelette. Cent ans plus tard, ces chiffres étaient ramenés à des valeurs allant de 0,01 à 1 % du squelette. De telles fluctuations dans les valeurs estimées sont principalement dues à la présence d'algues endolithiques, qui abondent dans les squelettes coralliens. Ces dernières peuvent induire une surestimation des valeurs, mais aussi une sous-estimation, en induisant une dégradation biologique de la MO. Au final, des valeurs plus précises ont été obtenues dernièrement par thermogravimétrie, et révèlent une valeur de 1 % pour la MO déshydratée, et 2,5 % pour la MO hydratée. Ces chiffres peuvent cependant être sous-estimés par des pertes successives, notamment des molécules de faibles poids moléculaires, lors des très nombreuses étapes de préparation des échantillons, mais aussi car une partie de la MO est insoluble, et se retrouve éliminée lors des étapes de préparation.
7.2. Synthèse de la MO
Dès 1903, des chercheurs décrivaient la synthèse et la sécrétion de molécules organiques au niveau des tissus squelettogéniques des coraux. Observations qui ont, par la suite, été confirmées à de nombreuses reprises par microscopie. Des méthodes d'immunohistochimie (détection et visualisation spécifique d'une ou d'un groupe de molécule choisi) ont montré récemment que seul l'ectoderme calicoblastique et plus précisément les calicoblastes étaient responsables de la synthèse et de la sécrétion par exocytose de la MO. Ces observations ont exclu l'hypothèse géochimiste, selon laquelle la MO serait des restes de tissus emprisonnés lors de la précipitation du CaCO3.
7.4. Caractérisation biochimique de la MO
Différentes familles de molécules ont été rapportées comme étant des composants de la MO. Ainsi, et selon les espèces, la MO pourrait contenir de la chitine et des substances de type hyaluronique (polysaccharide, c.à.d. un enchainement de molécule de sucre), des lipides et des protéines. Cependant, la plupart des articles révèlent la présence de protéines qui peuvent être sulfatées ou glycolisées, mais aussi de sucres simples ou complexes, sulfatés ou non. Le ratio entre protéines et sucres est très variable selon les espèces. Par exemple, chez Stylophora pistillata et Pavona cactus, la concentration en protéine est de l'ordre de 0,013 à 0,021 % du squelette, alors que la quantité de sucre est 17 fois plus importante dans la MO de S. pistillata que dans celle de P. cactus. Outre ces molécules plus ou moins complexes, la MO est également composée d'eau, ce qui suggère qu'il pourrait s'agir d'un hydrogel, dont le taux d'hydratation est très variable d'une espèce à l'autre.
D'un point de vu biochimique, une caractéristique majeure des protéines de la MO est leur extrême richesse en acides aminés acides, tel que l'acide aspartique. C’est d'ailleurs cette forte acidité qui rend ces protéines très difficiles à caractériser. La première protéine de MO à être caractérisée fut la galaxine du corail Galaxea fascicularis. Courte, glycosilée, ne fixant pas le calcium, elle est caractérisée par un enchainement d'acides aminés répétés en tandem. Trois protéines présentant les mêmes caractéristiques ont été découvertes récemment chez Acropora millepora. De très nombreuses protéines de MO restent cependant à découvrir et surtout à caractériser biochimiquement afin de mieux comprendre leurs fonctions. La partie sucrée de la MO est composée de sucres simples (monosaccharide), ou assemblés entre eux sous forme de glycosaminoglycan.
7.5. Le rôle de la MO dans la calcification
Il a tout d'abord été proposé que la MO était le facteur limitant de la calcification. Ainsi, la vitesse de production et de sécrétion de la MO dicteraient la vitesse maximum que pourrait prendre la calcification et donc la croissance dans son ensemble. En étudiant la répartition de la MO dans le squelette, certains auteurs ont proposé qu'elle pourrait jouer le rôle d'inhibiteur de la calcification, créant ainsi le moule qui donnerait sa forme au squelette. Plus récemment, il a été démontré que la calcification avait lieu en deux étapes. Tout d'abord synthèse et déposition de la MO au sein de laquelle prendrait naissance, dans un même temps, le centre de calcification et la fibre squelettique, formant ainsi le cristal dans son ensemble.
Parce que les protéines de la MO sont extrêmement riches en acides aminés acides, notamment sous forme de chaines d'acides aspartiques, il a été suggéré que la MO pouvait fixer du Ca2+ en grande quantité, mais avec une faible affinité (fixation peu résistante), et de façon réversible (possibilité de décrocher les ions Ca2+ liés à la MO). Ces capacités permettraient à la MO de concentrer, puis de libérer du Ca2+ quand ce dernier est nécessaire à la calcification. Puisque la fixation de Ca2+ a également été caractérisée au niveau des phopholipides du squelette, il a également été proposé que ces derniers pouvaient servir de base à la nucléation du CaCO3. Pour en revenir aux Galaxines, leurs rôles restent, encore aujourd'hui, énigmatiques. Cependant, très riches en acides aminés de type cystéines (deux cystéines peuvent se lier entre elles et former des ponts intramoléculaires), il a été proposé que les galaxines pouvaient être imbriquées les unes dans les autres et former ainsi un réseau très serré et complexe, offrant un support au reste des molécules de la MO. À ce jour, une enzyme particulière a été découverte dans la MO, l'anhydrase carbonique. Enzyme qui a la capacité de transformer du CO2 en CO32-, et donc de transformer le carbone inorganique dans sa forme utile à la calcification. Pour finir, il est aujourd'hui bien admis que la MO serait responsable de la morphologie du squelette et des éléments du squelette, étant donné que quantité, structure et composition de la MO, sont spécifiques à chaque espèce de corail. Tout comme la morphologie du squelette. Il apparait d'ailleurs que la MO est le négatif du squelette décalcifié que ce soit à l'échelle macroscopique ou microscopique
8. Le coût de la calcification
Il n'existe aucune donnée sur le coût énergétique de la calcification chez les coraux. Cependant, de nombreux faisceaux d'arguments sous-entendent un coût très important de cette fonction. Par exemple le transport d'ion est connu pour demander beaucoup d'énergie. Ainsi, ne serait-ce que pour assurer les échanges journaliers en K+/Na+, une cellule (quelle qu’elle soit) utiliserait 6 à 40 % de l'énergie qu’elle a à disposition. Chez les Coccolithophoridae (organisme calcifiant) le transport du Ca2+ représenterait une part majeure du coût total de la calcification. Chez les mollusques, il a été estimé que la calcification utilisait 7,5 fois plus d'énergie que la croissance tissulaire, et 17,5 fois plus que la reproduction. Parmi ces coûts énergétiques, 22 % seraient liés à la synthèse et la sécrétion de la MO, alors qu'elle ne représente que 1,5 % du poids de la coquille. Basé sur ces quelques chiffres, en partant du postulat que le squelette corallien était composé de 0,1 à 1 % de MO, et des mesures du budget énergétique total connu pour Porites porites, il a été évalué que la calcification coutait 13 à 30 % de l'énergie produite par un corail. La croissance tissulaire ne représentant que 8% de cette énergie
9. Conclusion
Ces quelques lignes ont permis d'établir les bases physiologiques et moléculaires de ce qu'est la calcification des coraux. Phénomène complexe et plurifactoriel, il reste aujourd'hui méconnu, même si de nombreuses avancées nous permettent d'y voir plus clair. Pour nous aquariophiles, de telles données peuvent ouvrir de nombreuses perspectives de maintenances. En effet, si 15 à 20 ans en arrière, maintenir un Acropora sp., et peut être le voir grandir de 2 à 3 mm par mois constituaient le graal à atteindre, aujourd'hui l'heure est aux bacs à boutures et à la production industrielle. Cependant, produire nécessite de comprendre, et comprendre permet d'optimiser, tout du moins d'essayer…
Compte tenu des données présentées, il semble que maintenir des paramètres de KH et de concentration en Ca2+ optimales permettraient une croissance plus rapide et continue. Un pléonasme me direz-vous, cependant si le problème est vu dans l'autre sens, on peut s'interroger sur l'importance de l'alimentation externe par voie hétérotrophe en condition non-optimale. En effet, très couteuse en énergie, il se pourrait que la calcification puisse tout de même être "maximale", si suffisamment d'énergie était disponible pour cette dernière. Si l'alimentation peut être un soutien essentiel de la croissance, la qualité des apports peut également être éclairée par les données présentées. En effet, nous avons pu voir ici le rôle de la matrice organique, véritable ossature du squelette. Cette dernière est un prérequis essentiel à la déposition du carbonate de calcium. Essentielle, mais particulière dans sa composition, la MO pourrait elle aussi être soutenue par des apports externes. En effet, quelque AA, clefs de cette MO, pourraient être apportés de façon plus spécifique afin d'en faciliter la synthèse. Citons notamment l'acide aspartique constituant majeur de certaines protéines de matrices, mais aussi les cystéines et les prolines, deux AA clefs dans la structuration des protéines, et donc du réseau moléculaire de cette fameuse ossature qu'est la MO.
Comme tout phénomène biologique, la calcification est indirectement en lien avec les facteurs externes tel que la température, le pH, ou encore la lumière... C'est dans une deuxième partie que ces interactions, et surtout leurs effets sur la calcification seront abordés.
Jérémie Vidal - Dupiol
Article publié par Cap Récifal le 21 juin 2013 avec l'aimable autorisation de l'auteur.
Commentaires recommandés
Il n’y a aucun commentaire à afficher.