

Concernant les poissons clowns (Amphiprion percula) le couple pond toujours à proximité d’une anémone qui correspond à leur lieu de vie. L’éclosion a lieu la nuit et les larves sont disséminées dans l’océan en fonction des courants et de leurs capacités natatoires. À la suite de cette phase océanique, qui dure une bonne dizaine de jours les larves retournent vers le récif et vont coloniser le lagon. C’est au cours de cette période qu’a lieu la métamorphose. Cette transformation correspond à une évolution ontogénétique et se traduit par un changement brutal de la morphologie, de la physiologie et du comportement des alevins. Durant cette période, les larves vont choisir un nouvel habitat qui va influencer directement leur survie et leur croissance ce qui correspond généralement à la phase d’installation (sédentarisation). Les juvéniles vont s’intégrer à des populations adultes : c’est la phase dite de recrutement.
1. Publication
Ces caractéristiques ont conduit à la création d’une multitude de niches écologiques permettant d'utiliser les ressources disponibles de façon optimale. L’image d’une oasis dans le désert fait également référence à la forte productivité de cet écosystème. Cependant la production dite nette (ce qui peut être simplifié par "ce qui est rejeté et mesurable dans le milieu") est proche de zéro, malgré une croissance forte (production) des producteurs primaires que sont les algues et les coraux.

De nombreux auteurs se sont interrogés sur la capacité des larves à détecter les récifs et à les coloniser ainsi qu’à identifier l’habitat idéal lors de la phase d’installation. Les larves sont susceptibles de percevoir les signaux environnementaux pour trouver un habitat grâce à leurs sens tels que l’olfaction, l’ouïe et la vue.
C’est dans ce cadre que s’inscrit cette publication Les poissons coralliens sentent des feuilles pour trouver leur îles natales, parue dans The Royal Society en 2008 : Coral reef fish smell leaves to find island homes (Danielle L. Dixson et al.)
2. Objectifs de l'étude
Les poissons marins commencent leur vie sous forme de larves qui vont se disperser dans l’océan. La capacité de trouver un habitat convenable après une période en eau libre représente un défi important pour les larves marines.
La théorie suggère que la persistance des populations de poissons de récifs coralliens nécessite non seulement une connectivité significative entre les sous-populations, mais aussi que certains jeunes retournent à des sous-populations natales. Les recherches récentes ont confirmé un niveau élevé d’auto-recrutement ainsi qu’une connectivité entre les habitats des récifs isolés. Cependant, les moyens par lesquels les larves des poissons coralliens détectent, s’orientent et s’installent sur les récifs restent un mystère.
Les progrès récents, dans notre compréhension des larves de poissons coralliens, indiquent qu’elles disposent de capacités sensorielles et de natation bien développées. Les larves de poissons coralliens peuvent utiliser les signaux olfactifs pour identifier les sites appropriés, une fois qu’ils sont dans le voisinage du récif. Cependant, les signaux chimiques que les larves utilisent pour trouver un habitat convenable restent mal compris.
Des travaux récents en Papouasie-Nouvelle-Guinée ont montré qu’une forte proportion de poissons-clowns juvéniles est recrutée à partir de leurs populations natales. L’objectif général de cette étude est d’étudier le rôle des signaux olfactifs potentiellement utilisés par les larves des poissons clowns (Amphiprion percula) pour localiser les foyers de l’île.
Tout d’abord, les auteurs ont examiné la distance à laquelle cette espèce se limite par rapport aux récifs frangeants des îles de la baie de Kimbe (Papouasie-Nouvelle-Guinée). Les auteurs ont ensuite effectué une série d’expérience sur le terrain, en utilisant un système de « canaux de choix », développé par Gerlach et al (2007) pour tester la capacité des juvéniles à différencier les odeurs générées par les îles environnantes. Les auteurs émettent l’hypothèse que les juvéniles perçoivent les signaux chimiques des feuilles des arbres tropicaux ainsi que des anémones pour retourner dans leur habitat natal. Enfin, pour vérifier si les larves sont influencées par l’habitat natal ou si elles présentent des préférences innées, ils ont également étudié des larves élevées en laboratoire, sans contact avec des anémones et la végétation.
3. Résultats
3.1. Association entre les récifs situés à proximités des îles tropicales
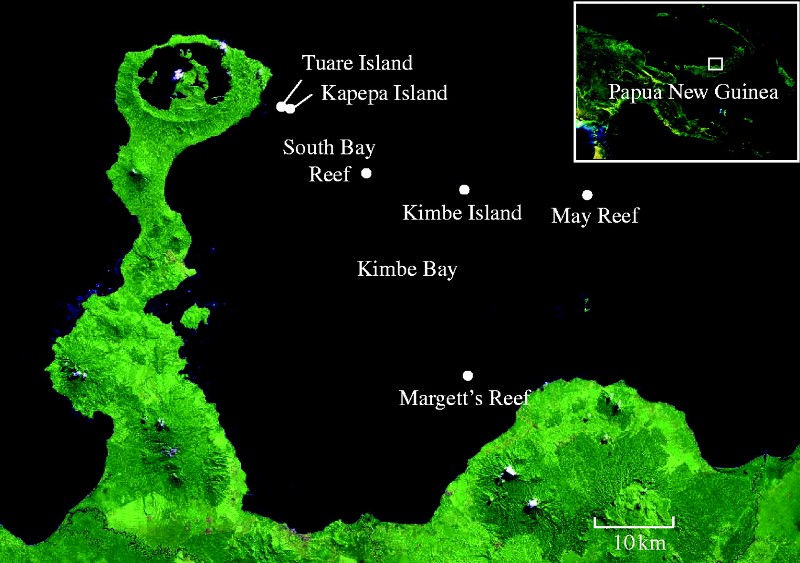
Les auteurs ont réalisé des comptages d’A. percula ainsi que de leurs anémones (Heteractis magnifica et Stichodactyla haddoni) sur 6 sites d’études à l’ouest de l’île de Kimbe Bay. À proximité de trois récifs, on constate la présence de petites îles couvertes de végétation tropicale ; sur les trois autres récifs il n’y a pas d’îles environnantes. Les résultats ont montré qu’A. percula et les anémones qu’ils occupent, présentent une forte association avec les récifs entourés d’îles. Les anémones et A. percula ont été rarement observés sur les récifs émergents sans présence d’îles à proximité.
Des « transects » visuels (ligne virtuelle ou réelle que l’on suit afin de réaliser un échantillonnage pour étudier un phénomène particulier) ont été réalisés afin d’évaluer l’abondance des feuilles des arbres tropicaux dans l’eau sur chacun des 6 sites et seuls les sites à proximité des îles présentaient des feuilles. De plus, sur deux récifs situés à proximité de deux îles les feuilles ont été comptées à différentes distances du rivage (moins de 5 m du rivage, crête récifale, 500 m en mer et à 1 km au large). Des feuilles flottantes ont été observées dans la première zone à proximité de la plage et le nombre diminue sensiblement avec la distance pour être nul à 1 km des côtes.
Une carte détaillée de l’emplacement de deux espèces d’anémones a été réalisée afin de visualiser leur répartition entre les différents types d’habitats. Les résultats ont montré qu’elles ne sont pas distribuées de façon aléatoire : S. haddoni a été presque toujours située à 10 m du rivage sous la végétation en surplomb et à proximité de la litière des feuilles. H. magnifica est plus communément présente, entre 10 et 30m du rivage dans des petites lagunes mais pas sur les zones importantes du platier ou sur la pente récifale.
3.2. Tests de choix olfactifs
Pour évaluer la capacité des juvéniles d’A. percula de choisir entre l’eau contenant potentiellement différents stimuli d’odeurs, les auteurs ont utilisé une boite de choix à travers deux canaux (13 cm × 4 cm) développée par Gerlach et al. (2007). Cet appareil a été conçu pour mener des expériences de choix afin que les poissons puissent s’orienter librement entre l’eau qui coule de deux sources différentes. L’eau des deux sources différentes est alimentée par gravité à partir de seaux et s’écoule au travers de tubes. Les poissons sont relâchés à l’extrémité avale après une période d’acclimatation de 5 minutes où ils sont libres de se déplacer vers la source d’eau préférée. Les juvéniles A. percula ont été capturés dans le milieu naturel auprès des anémones.
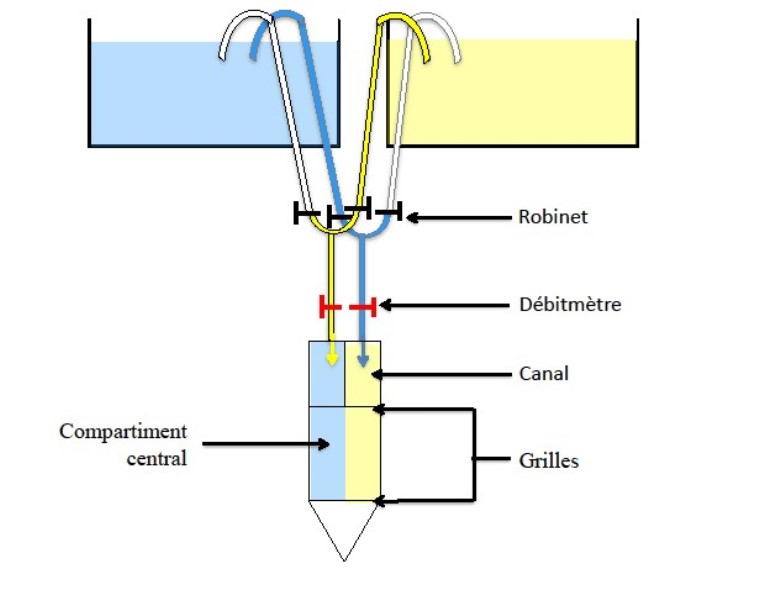

Des expériences de choix ont été conduites afin de voir si les jeunes alevins pouvaient différencier l’eau collectée dans les récifs situés à proximité des récifs où des îles ne sont pas présentes. A. percula présente une forte préférence pour les échantillons d’eau prélevés dans les récifs situés à proximité des îles.
3.2.1. Discrimination olfactive entre les échantillons d’eau prélevés à des distances différentes des récifs situés à proximité des îles
Une nouvelle série d’expériences de « tests de choix olfactif » a été réalisée en prélevant de l’eau à différentes distances des îles : eau prélevée à moins de 1 m de la côte, eau prélevée à proximité du bord extérieur de la crête du récif, et de l’eau prélevée à 1 km de l’île.
Pour les deux récifs insulaires étudiés, les juvéniles d’A. percula ont montré une forte préférence pour l’eau prélevée à proximité des plages, indépendamment du fait que les juvéniles sont confrontés à l’eau de la crête récifale ou à l’eau du large. Toutefois, les juvéniles n’ont montré aucune discrimination significative entre l'eau prélevée en mer et celle de la crête récifale.
3.2.2. Discrimination olfactive entre les échantillons d’eau traitée et non traitée avec l’anémone S. haddoni ou avec les feuilles des arbres de la forêt tropicale surplombant l’eau
Une nouvelle série d’expériences a été réalisée avec de l’eau prélevée au large mais traitée avec une anémone ou avec des feuilles de la forêt tropicale.
La réponse potentielle à des signaux chimiques de feuilles a été testée en ajoutant des feuilles déchirées de cinq arbres d’une île proche d’un récif. Les feuilles provenant des cinq espèces différentes ont été testées individuellement. Les résultats ont montré que les juvéniles étaient attirés par l’eau traitée avec l’anémone S. haddoni lorsqu’ils avaient le choix entre l’eau traitée et non traitée.
Il en est de même pour l’eau traitée avec des feuilles de la forêt tropicale aussi bien pour les cinq plantes prises individuellement que pour le lot des cinq feuilles.
3.2.3. Discrimination olfactive des larves élevées en laboratoire.
Afin de voir si les préférences olfactives observées sur les juvéniles capturés sur le récif sont acquises ou innées, des larves élevées en aquarium sans exposition préalable aux stimuli, ont été soumises à des expériences avec la même boîte à choix.
Pour tester si les juvéniles pouvaient discriminer les échantillons d’eau traitée avec des feuilles et avec leur anémone, les larves de différentes pontes ont été élevées pendant 11 jours. Les larves ont été testées avec différentes sources d’eaux traitées et non traitées avec une anémone.
Pour tester les réponses aux feuilles, les échantillons d’eaux ont été traités avec les feuilles d’un arbre de la forêt tropicale côtière commune (Chrysanthus xanthostemon : Myrtaceae).
Les larves d’élevage ont été fortement attirées par des signaux chimiques de l’anémone, indiquant qu’à partir du 11ème jour, elles sont capables d’avoir une réponse olfactive.
Les larves d’élevage ont une préférence également pour l’eau traitée avec des feuilles de l’arbre tropical par rapport à celle non traitée.
4. Discussion
Le poisson-clown A. percula a clairement une association étroite avec les récifs coralliens qui sont situés à proximité des îles ayant une forêt tropicale. Compte tenu de la forte association entre les anémones et les récifs situés à proximité des îles, les larves d’A. percula maximisent leurs chances de trouver un site approprié en étant capable de localiser et de s’orienter vers des îles. Les données expérimentales présentées ici suggèrent fortement que A. percula a une attraction olfactive innée pour la végétation de la forêt tropicale.
Du fait de la difficulté potentielle de trouver l’habitat de récif de corail après une phase larvaire pélagique, il est probable que A. percula utilise une suite de signaux sensoriels pour trouver les récifs de l’île, qui permettraient aux larves, par la suite, de trouver un habitat convenable.
Les fortes densités d’anémones et d’A. percula adultes sur les récifs situés à proximité des îles font certainement partie du cocktail de sensations utilisé pour trouver un endroit où s’installer. Les poissons de récif sont capables de répondre aux signaux chimiques des anémones mais également de leurs congénères.
Cependant, les signaux olfactifs provenant de ces sources sont plus importants une fois que les larves sont à proximité de récifs. Il est probable que les signaux terrestres peuvent être détectés à des distances plus grandes, étant donné que les feuilles et les débris de végétaux flottent sur des distances considérables.
Les niveaux élevés d’auto-recrutement sur les récifs situés à proximité d’une île soulèvent d’autres questions, notamment sur la capacité des larves à trouver des foyers insulaires. Par exemple, Almany et al. (2007) ont montré que, après une période de 10 à 12 jours planctoniques, jusqu'à 60 % des juvéniles A. percula installés sur un récif d’une île étaient les descendants de résidents adultes. La manière dont ces jeunes ont trouvé leur chemin de retour à l’île est inconnue, mais leur capacité à réagir à des stimuli olfactifs des îles est clairement exploitée.
Il est possible que les larves d’A. percula soient attirées sur leur île natale en réponse à des indices de végétation spécifiques. Même si les larves ont une capacité innée à répondre à la végétation de la forêt tropicale, elles peuvent aussi être capables de percevoir des signaux chimiques rencontrés pendant le stade embryonnaire.
Il est indispensable d’étudier à quelle distance les larves d’A. percula sont capables de détecter les signaux chimiques et à quel stade du développement elles peuvent y répondre. De plus, les produits responsables de cette attraction n’ont pas été identifiés mais les tanins et des composés voisins des tanins, issus d'algues et d'herbiers, sont souvent impliqués dans l’attraction des larves de nombreux organismes marins benthiques.
Les résultats de cette étude ont des répercussions importantes dans la gestion des écosystèmes des îles proches des récifs. A. percula est un exemple du lien fort qui existe entre les récifs coralliens et la forêt tropicale environnante. Dans les environnements insulaires, la gestion intégrée des habitats terrestres et marins est essentielle. La déforestation et le développement des îles pourraient avoir un impact négatif sur la capacité des poissons clowns à trouver un habitat convenable.
5. Conclusion
Une larve est-elle amenée sur le récif, telle une particule passive ? Son arrivée dépend-elle d’une décision, c’est-à-dire d’une réaction de la larve face à son environnement ? Les comportements complexes décrits précédemment, ajoutés aux capacités natatoires des larves, particulièrement celles développées aux stades de postflexion et de transition, laissent supposer que la colonisation est un processus actif.
Cette étude montre que les larves peuvent s’orienter vers des récifs spécifiques, même voisins d’autres récifs, car elles détectent des odeurs différentes. Elles sont capables d’identifier l’odeur du récif natal, ce qui induit une limitation de la dispersion. Le fait de retourner chez soi est la première étape de l’isolement des reproducteurs, qui pourrait mener à la spéciation (processus évolutif de développement d’une nouvelle espèce). En effet cela souligne l’isolement des populations (populations localisées en un lieu donné) et provoquerait la divergence génétique des populations.
Ces études suggèrent que ces populations sont constituées, en fait, par des réseaux de populations connectés ensemble formant une métapopulation (ensemble de population d’individus d’une même espèce séparé géographiquement). Il reste à découvrir la variabilité temporelle et l’intensité de ces connexions.
6. Pour aller plus loin…
Pourquoi réaliser une phase pélagique dans ce cas ?
Cette stratégie est suivie par de nombreux poissons coralliens, voire de nombreux organismes benthiques, car elle apporte beaucoup d’avantages. La stratégie de dispersion larvaire dans le milieu océanique permettrait d’éviter la forte prédation présente sur les récifs coralliens (zooplanctonophage). Le développement larvaire dans l’océan, réclame également une dépense d’énergie moindre par les larves, par rapport à celle nécessaire pour lutter contre un hydrodynamisme violent dans cette zone. De plus, en exploitant les ressources du milieu pélagique, les larves ne sont pas en compétition avec les adultes. Enfin, cette stratégie d’étalement dans un environnement fragmenté, permet de limiter les risques. Les populations locales sont soumises à des perturbations à des échelles de temps différentes (milliers d’années par rapport à la variation du milieu marin) ou à des échelles plus courtes (développement d’un prédateur, phénomènes climatiques, pollution…) qui peuvent engendrer des destructions totales de l’habitat et de l’espèce. Cette stratégie de dispersion larvaire serait une probable adaptation aux fluctuations des facteurs environnementaux et à l’instabilité de l’écosystème corallien. Dans le cycle biologique, la phase de dispersion pélagique se caractérise par un taux de mortalité très élevé que l’on suppose proche de 99 % pour les poissons coralliens.
Puis, vient l’étape de colonisation, de courte durée. C’est une étape importante pour les larves qui passent du milieu océanique au milieu récifal (variations de température, de salinité). Pour un certain nombre de poissons, c’est à cette étape qu’a lieu une métamorphose morphologique et physiologique des larves.
C’est lors de la phase d’installation que les juvéniles doivent trouver un habitat. Cette étape est aussi marquée par une forte mortalité. La recherche d’un habitat est conditionnée par la disponibilité des ressources alimentaires, par les populations déjà installées (compétition, prédation…) et par la présence d’un habitat adapté (dimensions, complexité, nature…). Là encore, les récifs coralliens offrent un milieu très propice à l’installation des juvéniles. La richesse trophique qui caractérise les récifs ainsi que la diversité des habitats qu’ils peuvent proposer sont autant d’avantages pour les juvéniles.
Le suivi de la colonisation et de l’installation des larves permet de comprendre et même de prédire en partie la dynamique des populations adultes par la suite. Ainsi, les larves des poissons coralliens seraient capables de détecter et de localiser les informations émises par les « conspécifiques » (individus de la même espèce) et par leurs habitats.
Les larves disposent de capacités natatoires suffisantes pour retrouver leur habitat une fois qu’elles l’ont détecté. C’est grâce à leurs sens qu’elles arrivent à localiser les agrégats d’habitats sur lesquels elles peuvent s’installer. Ainsi l’ouïe permettrait aux larves de détecter un récif. La vision et la ligne latérale seraient utilisées pour éviter les prédateurs. Enfin, à proximité du récif, l’odorat favorisait l’installation lors de la localisation de l’habitat et la vision serait utilisée lors de la phase d’installation.
7. Encore un peu plus loin… Nature versus Nurture ou l’inné face à l’acquis
Quelle est la part des comportements qui sont déterminés par la génétique ? Par opposition, quelle part est acquise lors de l’interaction de l’individu avec son environnement ? Cette question entraîne bien souvent des prises de positions passionnées et je ne me risquerais pas à trancher.
Cette publication nous apporte une explication sur l’attraction d’A. percula pour son anémone. En effet, bien qu’il soit d’élevage et qu’il ne connaisse pas son futur hôte : H. magnifica, le poisson-clown est irrémédiablement attiré par son anémone. Génétiquement, le poisson-clown est destiné à rencontrer son anémone.
En effet, il n’est pas sculpté pour la nage, de plus il a besoin de dormir profondément. Cela fait de lui une cible facile s’il n’avait pas eu la sage décision de se faire protéger par un hôte urticant dans le milieu naturel. En aquarium en l’absence d’anémone, il trouve souvent un hôte de substitution.
Évidemment, cela demande des études complémentaires pour identifier la ou les molécules chimiques qu’il arrive à percevoir. On peut également émettre l’hypothèse que chaque anémone dispose d’un arsenal chimique différent qui attire plus ou moins spécifiquement une ou plusieurs espèces de poissons-clowns. C’est pourquoi certains d’entre nous n’arrivent pas à faire coexister leur anémone avec des poissons-clowns issus de régions géographiques totalement différentes.
Dans ce cas, à quand, pour nous aquariophile, un spray contenant les molécules chimiques attractives qu’il suffirait de pulvériser sur l’anémone ou le corail hôte afin d’attirer les poissons-clowns à l’endroit où l’on veut ?
Références
- Almany G.R, Berumen M.L, Thorrold S.R, Planes S, Jones G.P2007. Local replenishment of coral reef fish population in a marine reserve. Science. 316, 742–744.
- Gerlach G, Atema J, Kingsford M.J, Black K.P, Miller-Sims V2007. Smelling home can prevent dispersal of reef fish larvae. Proc. Natl Acad. Sci. USA. 104, 858–863. doi:10.1073/pnas.0606777104
Olivier SOULAT
Article publié par Cap Récifal le 11 mars 2015 avec l'aimable autorisation de l'auteur.
Commentaires recommandés
Il n’y a aucun commentaire à afficher.