

1 : Classification, anatomie - 2 : Nutrition, reproduction - 3 : Écologie - 4 : Maintenance
1. Classification
1.1. Sous classe Octocorallia
Les Octocoralliaires appartiennent à l’embranchement des cnidaires ("animaux-orties" en allusion à leurs cnidocytes) et à la classe des anthozoaires ("animaux-fleurs" en rapport à leur symétrie radiale). La classification des Octocoralliaires a beaucoup évolué ces dernières années et il n’est pas rare d’être perdu dans des descriptions antérieures.
On assimile bien souvent les zoanthaires (Zoanthus, Palythoa, Isaurus…) et les corallimorphaires (Actinodiscus, Discosoma, Ricordea…), dont les formes sont molles et dépourvues de squelette, à ce que l'on a coutume de nommer "coraux mous". Et pourtant, tout comme les Antipathaires (coraux noirs) souples, qui ressemblent aux gorgones, ces espèces appartiennent à la sous-classe Hexacorallia qui inclut les "coraux durs".
De nombreux récifalistes nomment les Octocoralliaires sous le terme "coraux mous", désignant ainsi les coraux n’ayant pas de squelette massif ou axial (les alcyons) et en leur associant parfois les gorgones, avec leur tige axiale semi-rigide. Mous, certains Octocoralliaires ne le sont pourtant pas, comme le corail bleu Héliopora coerulea ou Tubipora spp. qui forment des colonies au squelette rigide. La notion de « corail mou » restera à jamais une nébuleuse et nous devrions définitivement éviter d'utiliser cette dénomination.
Les scientifiques ne sont malheureusement pas tout à fait d’accord sur le classement des Octocoralliaires, à en juger le Système d'information taxonomique intégré (SITI) et celui du World Register of Marine Species (WoRMS), comme le montre le tableau 1. Stolonifera est en effet classé en tant qu’ordre par SITI et sous-ordre par Worms de plus SITI reconnait l’ordre Telestacea alors que WoRMS l'ignore.
| Noms communs | Système d'information taxonomique intégré SITI |
World Register Of Marine Species WoRMS |
|---|---|---|
| Alcyoninés | Ordre : Alcyonacea Sous ordre : Alcyoniina |
Ordre : Alcyonacea Sous ordre : Alcyoniina |
| Gorgones | Ordre : Alcyonacea Sous ordre : Holaxonia Sous-ordre : Scleraxonia Sous-ordre : Calcaxonia |
Ordre : Alcyonacea Sous ordre : Holaxonia Sous-ordre : Scleraxonia Sous-ordre : Calcaxonia |
| Stolonifères | Ordre : Stolonifera | Ordre : Alcyonacea Sous-ordre : Stolonifera |
| Heliopora | Ordre : Helioporacea | Ordre : Helioporacea |
| Plumes de mer | Ordre : Pennatulacea | Ordre : Pennatulacea |
| Ordre : Telestacea | - |
Il n'y a pas si longtemps, les Alcyoninés et les gorgones (qui constituaient les coraux mous aux yeux des scientifiques) étaient répartis dans des ordres différents. Comme le montre le tableau ci-dessus, les taxinomistes modernes sont aujourd’hui d’accord pour les regrouper dans le même ordre Alcyonacea (Alcyonacés ou Alcyonaires). L'illustration 1 propose un classement courant de la sous-classe Octocorallia, suivant 3 ordres : Alcyonacea, Helioporacea et Pennatulacea.

1.2. Espèces
Il existe encore de nombreuses divergences quand il s'agit de rentrer dans le détail des nomenclatures et notamment dans le classement des genres au sein des familles. Le tableau 2 présente la sous-classe Octocorallia selon le WoRMS et donne un aperçu non exhaustif des genres concernés.
| Ordres | Sous-ordre | Familles | Spécimen (ph.) | Genres |
|---|---|---|---|---|
| Alcyonacea (Alcyonacés) (Alcyonaires) |
Alcyoniina (Alcyoninés) |
Alcyoniidae (Alcyoniidés) |
 
|
Acrophytum , Alcyonium, Aldersladum, Anthomastus, Bellonella, Cladiella, Dampia, Dimorphophyton, Discophyton, Drifa, Elbeenus, Eleutherobia (Photo 1), Heteropolypus Inflatocalyx, Klyxum, Lampophyton, Lanthanocephalus, Litophyton, Lobophytum, Lobularia Lohowia, Malacacanthus, Minabea, Notodysiferus, Paraminabea, Parerythropodium, Protodendron, Pseudoanthomastus, Rhytisma, Sarcophyton (Photo 2), Sinularia, Skamnarium, Sphaeralcyon, Thrombophyton, Verseveldtia |
| Nephtheidae (Nephteidés) |

|
Capnella, Chondronephthya, Chromonephthea, Coronephthya, Dendronephthya, Gersemi, Lemnalia, Leptophyton, Neospongodes, Nephthea, Pacifiphyton, Paralemnalia, Pseudodrifa, Scleronephthya, Stereonephthya, Umbellulifera (Photo) | ||
| Nidaliidae (Nidaliidés) |

|
Agaricoides, Chironephthya (Photo), Nephthyigorgia, Nidalia, Nidaliopsis, Orlikia, Pieterfaurea, Siphonogorgia | ||
| Paralcyoniidae (Paralcyoniidés) |
 |
Maasella (Photo) | ||
| Xeniidae (Xéniidés) |
 |
Anthelia, Asterospicularia, Cespitularia, Funginus, Efflatounaria, Heteroxenia, Sympodium, Sansibia, Xenia (Photo) | ||
| - | Acanthoaxiidae Haimeidae Paramuriceidae Parasphaerascleridae Viguieriotidae |
 |
Acanthoaxis - Villogorgia, Muriceides, Thesea Parasphaerasclera Studeriotes (Photo) |
|
| Holaxonia (Gorgones) |
Acanthogorgiidae |  |
Acanthogorgia (ex Acalycigorgia) (Photo), Anthogorgia, Calcigorgia, Muricella, Cyclomuricea | |
| Gorgoniidae |  |
Adelogorgia, Antillogorgia, Eugorgia, Eunicella, Filigorgia, Gorgonia, Guaiagorgia, Hicksonella, Leptogorgia, Lophogorgia, Olindogorgia, Pacifigorgia, Phycogorgia, Phyllogorgia, Pinnigorgia, Pseudopterogorgia (Photo), Pterogorgia, Rumphella, Tobagogorgia, | ||
| Plexauridae |  |
Astrogorgia, Bebryce, Caliacis, Echinomuricea, Echinogorgia, Eunicea, Euplexaura, Hypnogorgia, Lytreia, Menella , Muricea, Muriceides, Muriceopsis, Paracis, Paramuricea, Placogorgia, Paraplexaura (Photo), Plexaura, Plexaurella, Psammogorgia, Pseudoplexaura, Scleracis, Spinimuricea (ex Echinomuricea), Swiftia, Thesea, Trimuricea, Villogorgia | ||
| Keroeididae | Keroeides, | |||
| Calcaxonia (Gorgones) |
Ellisellidae |  |
Ctenocella, Dichotella, Ellisella, Heliania, Junceella (Photo), Nicella, Verrucella, Viminella, |
|
| Chrysogorgiidae Dendrobrachiidae Ifalukellidae Isididae Primnoidae |
 |
Stephanogorgia, Dendrobrachia Ifalukella, Plumigorgia, Isis, Jasminisis, Pteronisis, Zignisis, Calyptrophora (Photo), Callogorgia, Plumarella |
||
| Scleraxonia (Gorgones) |
Anthothelidae |  |
Alertigorgia, Anthopodium, Anthothela, Diodogorgia, Erythropodium, Homophyton, Iciligorgia, Solenocaulon (Photo), Semperina, Titanideum | |
| Briareidae Coralliidae |
 |
Briareum (Photo) Corallium, |
||
| Melithaeidae |  |
Acabaria (Photo), Clathraria, Melithaea, Mopsella, Wrightella, | ||
| Paragorgiidae Parisididae |
 |
Paragorgia (Photo) Parisis, |
||
| Subergorgiidae |  |
Subergorgia (Photo), Annella | ||
| Stolonifera (Stolonifères) |
Clavulariidae |  |
Carijoa, Clavularia (Photo), Sterosoma, Paratelesto, Telesto | |
| Acrossotidae Arulidae Coelogorgiidae Cornulariidae Pseudogorgiidae Tubiporidae |
 |
Acrossota Arula Coelogorgia, Cervera, Cornularia Pseudogorgia Tubipora (Photo) |
||
| Helioporacea | Helioporidae Lithotelestidae |
 |
Heliopora (Ph.) Epiphaxum |
|
| Pennatulacea (Pennatules) |
Sessiliflorae | Umbellulidae Veretillidae Protoptilidae ... |
 |
Umbellula, Cavernularia (Photo), Cavernulina, Veretillum... Distichoptilum, Protoptilum... ... |
| Subsessiliflorae | Halipteridae Pennatulidae Virgulariidae |
 |
Halipteris Crassophyllum , Pennatulata... Acanthoptilum, Virgularia (Photo)... |
|
| - | Chunellidae Echinoptilidae Pennautlidae Rennilidae Scleroptilidae Stachyptilidae |
 |
Chunella Actinoptilum, Echinoptilum - Renilla (Photo) Calibelemnon , Scleroptilum Gilibelemnon, Stachyptilum |
|
L’identification d'un spécimen n’est pas toujours aisée : son aspect peut varier en fonction des facteurs environnementaux, ainsi deux individus apparemment différents peuvent être des morphes de la même espèce. De plus, des coraux apparemment identiques peuvent appartenir à des espèces, à des genres, voire à des familles différentes. Bien souvent l’identification s’arrête au genre, notamment pour les Alcyonnaires. Comme on le verra, une identification formelle de l’espèce n’est possible que grâce à un examen microscopique des constituants de leur squelette interne, les sclérites ou spicules.
2. Anatomie des Octocoralliaires
2.1. Morphes
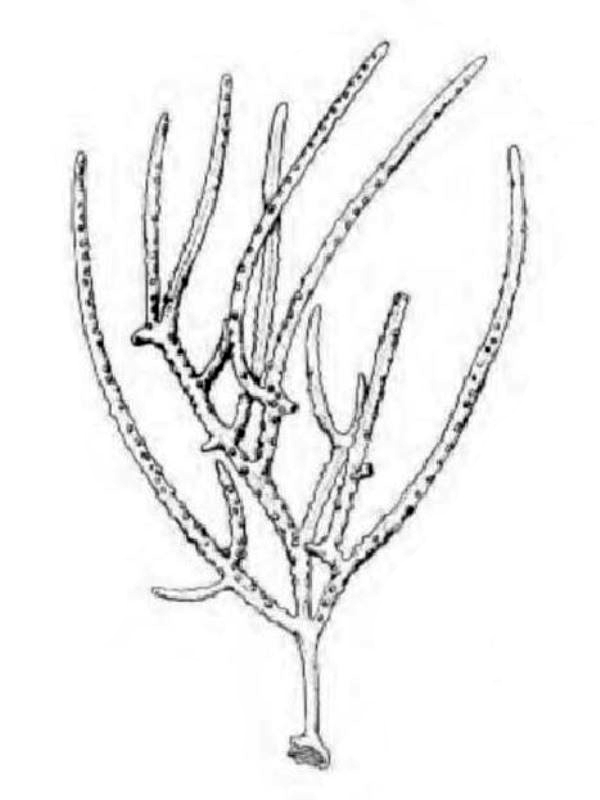
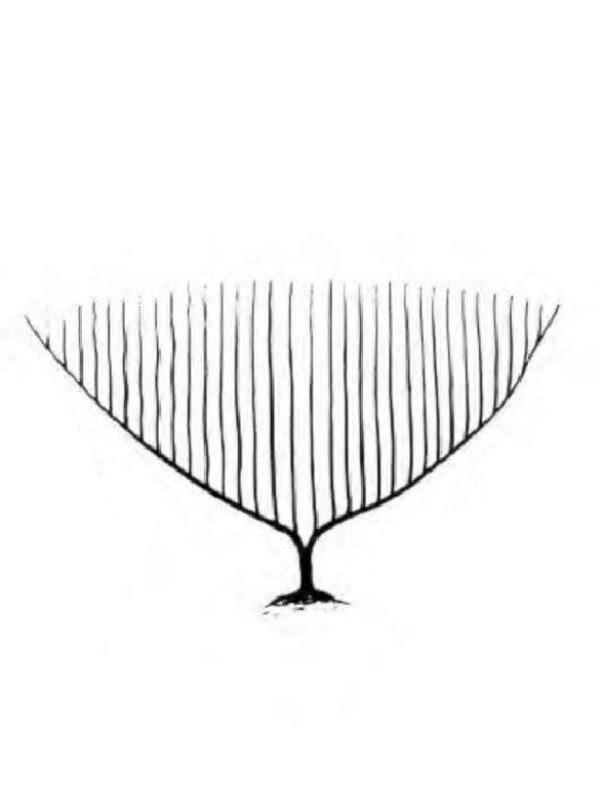
D'une manière générale, on distingue une portion basilaire (pied) surmontée d’une région plus ou moins ramifiée, portant les polypes (tiges, branches, lobes...), mais les formes peuvent varier notablement d'un genre à l'autre (illustration 2). Les Octocoralliaires sont le plus souvent constitués d'une colonie de polypes, il existe cependant des espèces monopolypes (solitaires) comme les Pennatules. Le pied est parfois bien distinct ou au contraire, il peut former une membrane (Briareum). Le capitule peut se déployer en ombrelle massive ou sous forme de dentelle légère.
Molles, souples ou parfois rigides, habituées du lagon ou des très grands fonds, des mers chaudes, tempérées et même froides... les espèces ne manquent pas pour qui veut s'intéresser aux Octocoralliaires.








"Ramifié", "arborescent", "digité"... beaucoup, trop de qualificatifs étaient employés pour définir ces formes souvent complexes. Les nombreux synonymes parfois sans correspondance linguisitique, ajoutaient à la difficulté de communiquer. Un groupe de biologistes a contribué à uniformiser, en plusieurs langues, le vocabulaire relatif aux Octocoralliaires. Ces travaux ont été publiés en 1983 par les professeurs Bayer, Grasshoff et Verseveldt dans le rapport Illustrated Trilingual Glossary of Morphological and Anatomical Terms Applied to Octocorallia. L'illustration 2 reprend une partie de cette terminologie relative aux morphes. Il n'est pas rare de devoir utiliser plusieurs termes pour caractériser un spécimen. Malheureusement, cette étape ne suffit pas à préciser le genre et encore moins l'espèce, comme on le verra plus loin.




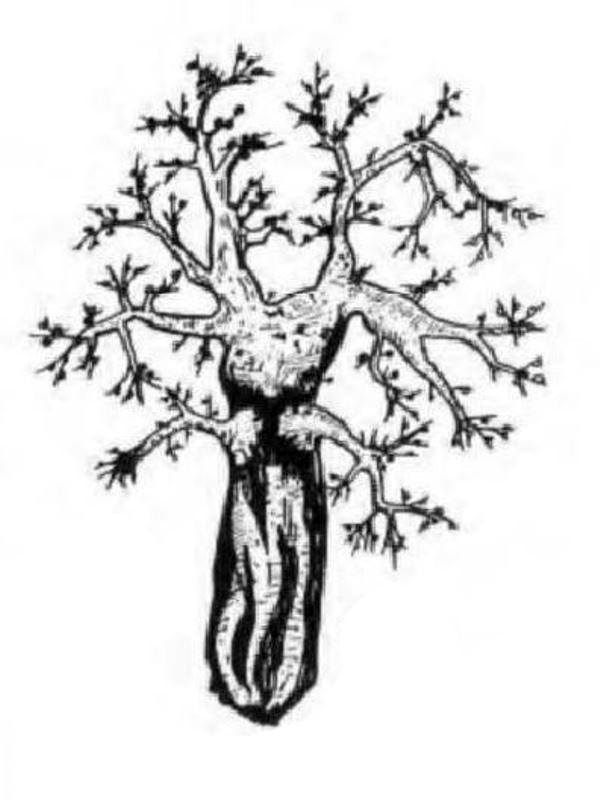


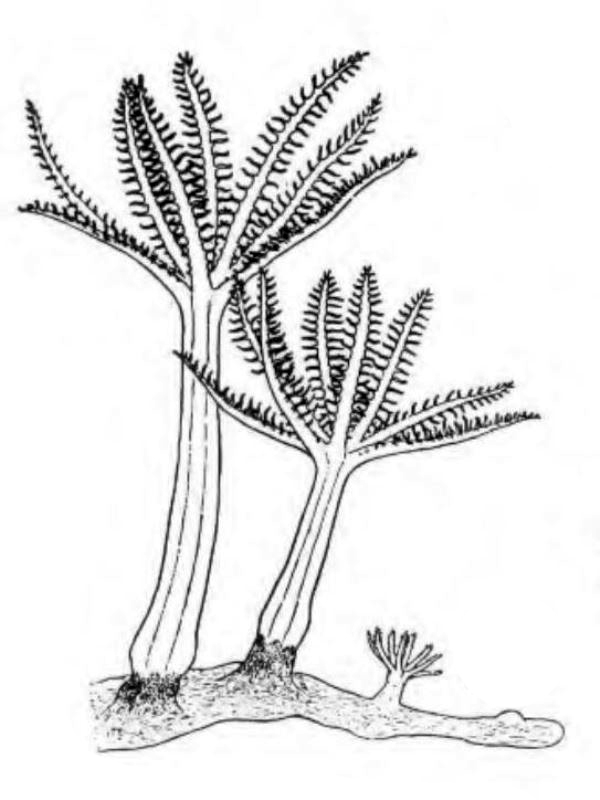
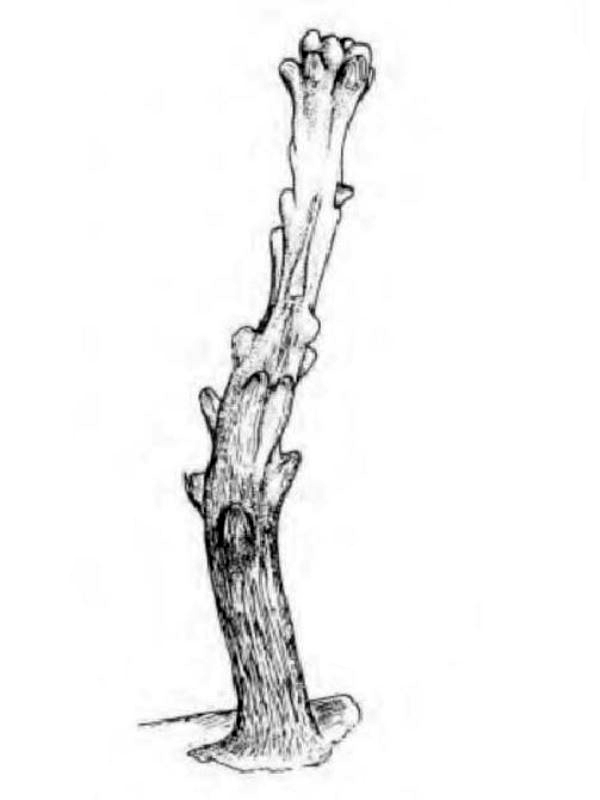


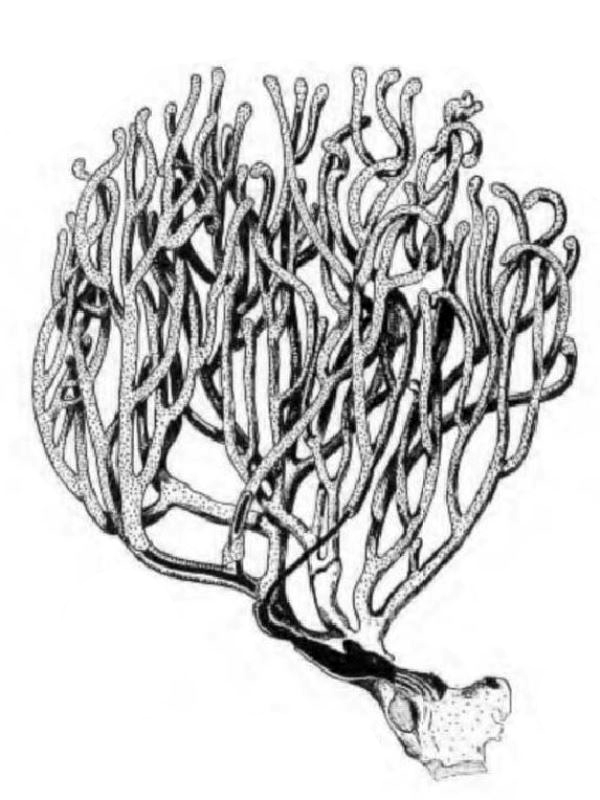








2.2 Polype

Les Octocoralliaires sont facilement reconnaissables : sauf rares exceptions, le polype présente, une symétrie d'ordre 8, ce qui se traduit extérieurement par la présence de huit tentacules et intérieurement, la cavité gastrique est divisée par huit cloisons. D’une manière générale, leurs tentacules possèdent des ramifications latérales en forme de plumes les pinnules (que n'ont pas les polypes de Scléractiniaires).
2.2.1. Couches cellulaires
Comme tous les Cnidaires, les Octocoralliaires sont des organismes diploblastiques c'est-à-dire qu'ils sont formés à partir de deux feuillets cellulaires, l'endoderme et l'ectoderme. Entre ces deux feuillets on trouve une matrice, la mésoglée.
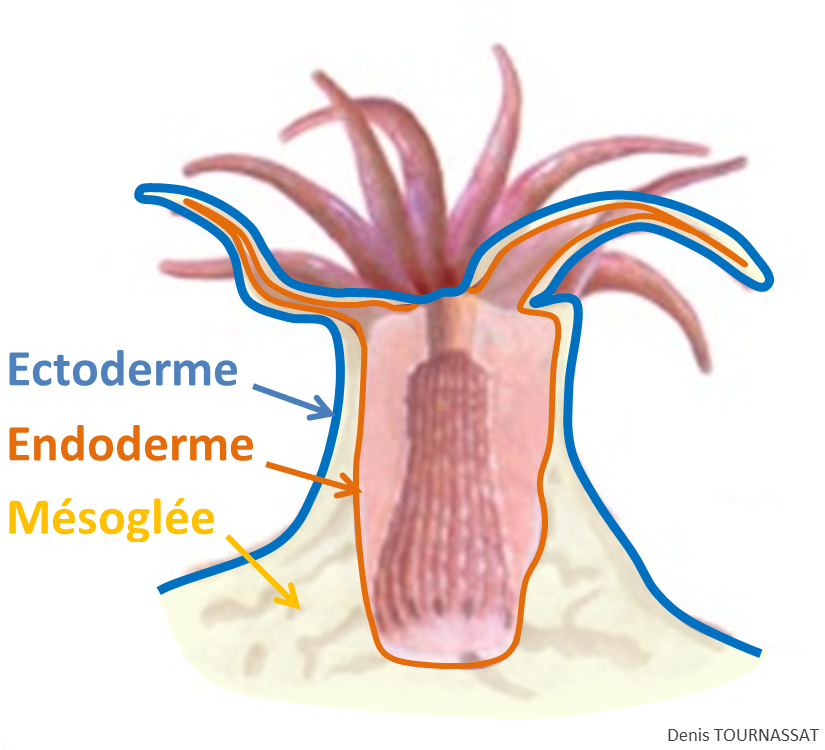
L’ectoderme : ce tissu externe couvre la surface du polype et de la colonie (le cœnenchyme). La zone couvrant le polype comporte des cellules sensorielles (détection), des cellules à mucus (captation et défense chimique), et des cnidocytes ou cnidoblastes (chasse et défense). La zone couvrant le cœnenchyme comporte moins de cellules sensorielles et de cnidocytes. On peut constater la faculté de certaines espèces à émettre une forte quantité de mucus, d’odeur caractéristique, destiné à protéger l’animal de la dessiccation lors des expositions à l’air durant les périodes d’exondation. Outre ce rôle, le mucus, riche en molécules antimicrobiennes, est aussi la première barrière immunitaire de ces organismes.
L’endoderme : ce tissu interne chargé de la digestion et de la reproduction, correspond à la couche interne des tentacules, de l’anthocodie (voir chapitre 2.3) et de la totalité des cavités internes. C'est dans les cellules de ce tissu que sont hébergées les zooxanthelles des espèces symbiotiques.
La mésoglée : c’est une matrice gélatineuse, située entre les deux couches ci-dessus. Elle ne constitue pas un véritable tissu puisqu'elle ne contient pas de cellules organisées (structurées) entre elles et donc par définition aucun organe différencié. D’épaisseur et de composition cellulaire variable selon l’espèce, elle est très fine au niveau des polypes mais représente la plus grande partie du cœnenchyme, entre ces derniers. Au niveau du cœnenchyme, la mésoglée abrite un réseau de canaux endodermiques (solenia) qui font communiquer toutes les cavités gastriques entre elles, répartissant ainsi les nutriments dans la colonie. Ce réseau est particulièrement développé chez les espèces massives comme Sarcophyton spp., il est réduit chez les espèces émergeant peu, à partir d’un tissu encroutant ou de stolons. Dans la mésoglée siège le système nerveux. Elle contient également les sclérites et des cellules chargées de la production du squelette.
2.2.2. Zooxanthelles et symbiose
Les zooxanthelles se développent en présence de lumière. Elles réalisent la photosynthèse qui consiste en la fabrication de composés carbonés à partir d’eau et de dioxyde de carbone sous l’action énergétique de la lumière.
Comme tous les végétaux photosynthétiques, les zooxanthelles contiennent des pigments qui leur confèrent une couleur brune, jaune ou verte : chlorophylles a et c, caroténoïdes et phycobilines. Ces pigments sont les molécules responsables soit directement de la photosynthèse, soit comme transmetteur de lumière, élargissant ainsi le spectre des longueurs d’onde utilisables. Ces pigments variés permettent aux zooxanthelles de s’adapter aux conditions d’éclairement (ensoleillement, profondeur…) et d’optimiser l’énergie lumineuse disponible. Ils présentent un pic d’absorption maximal situé entre 408 et 475 nm, c’est à dire dans la bande bleue extrême du spectre visible.
Le taux de photosynthèse croit avec l’intensité lumineuse, jusqu’à une valeur limite au-delà de laquelle il n’augmente pas. En situation d’éclairement extrême, notamment en présence d’UV (photo oxydation) leur densité diminue, elles dégénèrent et sont rejetées : les coraux blanchissent (phénomène de blanchissement des coraux). Cette situation est cependant réversible si les conditions redeviennent favorables.
Certaines espèces d’Octocoralliaires dites zooxanthellées, sont pourvues d’algues symbiotiques : les zooxanthelles, qui influent de manière notable dans leur métabolisme. Les Octocoralliaires dépourvus de ces algues sont dit azooxanthellés ou non photosynthétiques (NPS). Le tableau 2 détermine pour certains genres, leur caractère zooxanthellé ou azooxanthellé.
2.2.2.1. Les zooxanthelles

Ce sont des algues Dinoflagellés unicellulaires du genre Symbiodinium, de longueur environ 10 µm. Elles peuvent avoir une vie libre dans le milieu extérieur mais, concernant le corail, elles sont insérées, à raison d’une à trois cellules végétales, dans les cellules endodermiques de leur hôte. Lors de la reproduction sexuée, les zooxanthelles sont soit déjà présentes dans l'œuf du corail, soit ingérées en pleine eau aux stades larvaires ou juste après fixation et métamorphose. Lors de la reproduction asexuée, elles sont transmises via les tissus de la colonie mère. Les zooxanthelles se multiplient par mitose (division d'une cellule mère en deux cellules filles) au sein du corail et probablement aussi par voie sexuée hors du corail. Lors de la phase larvaire, leur population peut doubler en 10 jours. Cette biomasse végétale élevée représente alors une source d'énergie non négligeable.
Plus abondantes dans les parties superficielles exposées à la lumière : disque oral, cœnosarque et tentacules, leur densité et leur répartition dans les tissus varient selon plusieurs paramètres : les colonies de forme lamellaires développent bien naturellement plus d’algues sur leurs faces supérieures ; la densité varie sur la hauteur des espèces digitées ; les polypes en expansion le jour renferment d’avantage de zooxanthelles, notamment au niveau des tentacules ; la densité de ces algues symbiotiques augmente également avec la concentration en ions NH4+ dans l’eau. Mais la lumière semble bien être le facteur le plus important.
2.2.2.2. Symbiose

Il s'agit d'une association durable et profitable (symbiose mutualiste) entre deux organismes : le polype hébergeur (hôte) et les zooxanthelles hébergées (symbiote). L’algue étant incluse dans une vésicule intracellulaire (le symbiosome) du polype, on parle d’endosymbiose.
Les zooxanthelles bénéficient de leur hôte : elles trouvent au sein des tissus, un environnement stable, à l'abri de la sédimentation de la prédation et des infections ; elles utilisent les déchets issus du métabolisme du corail pour leur propre métabolisme : le carbone à partir du gaz carbonique, en complément de celui provenant des hydrogénocarbonates dissous dans l’eau de mer ; les composés phosphorés (phosphates), azotés (ions NH4) comme source d'éléments minéraux qui sont localement plus concentrés que dans le milieu extérieur et dans une moindre mesure, nitrites et nitrates ; l’acétate converti en acides gras.
En retour, outre l’élimination des déchets azotés et phosphorés du corail, les zooxanthelles jouent un rôle important dans son métabolisme. Elles produiraient du dioxygène O2 utilisé par le tissu animal et agiraient dans la calcification : le prélèvement du CO2 par les zooxanthelles favoriserait la précipitation du carbonate de calcium et donc l'élaboration du squelette. Elles contribuent également largement à la nutrition des coraux qui les hébergent.
2.3. Deux types de polypes : autozoïdes et siphonozoïdes
Les Octocoralliaires disposent de polypes autozoïdes (ou gastrozoïdes) émergeant du corps. Certaines espèces possèdent en plus des polypes syphonozoïdes dont on ne distingue que l’orifice buccal au ras de l’épiderme. Ces deux types de polypes ont des fonctions spécialisées.
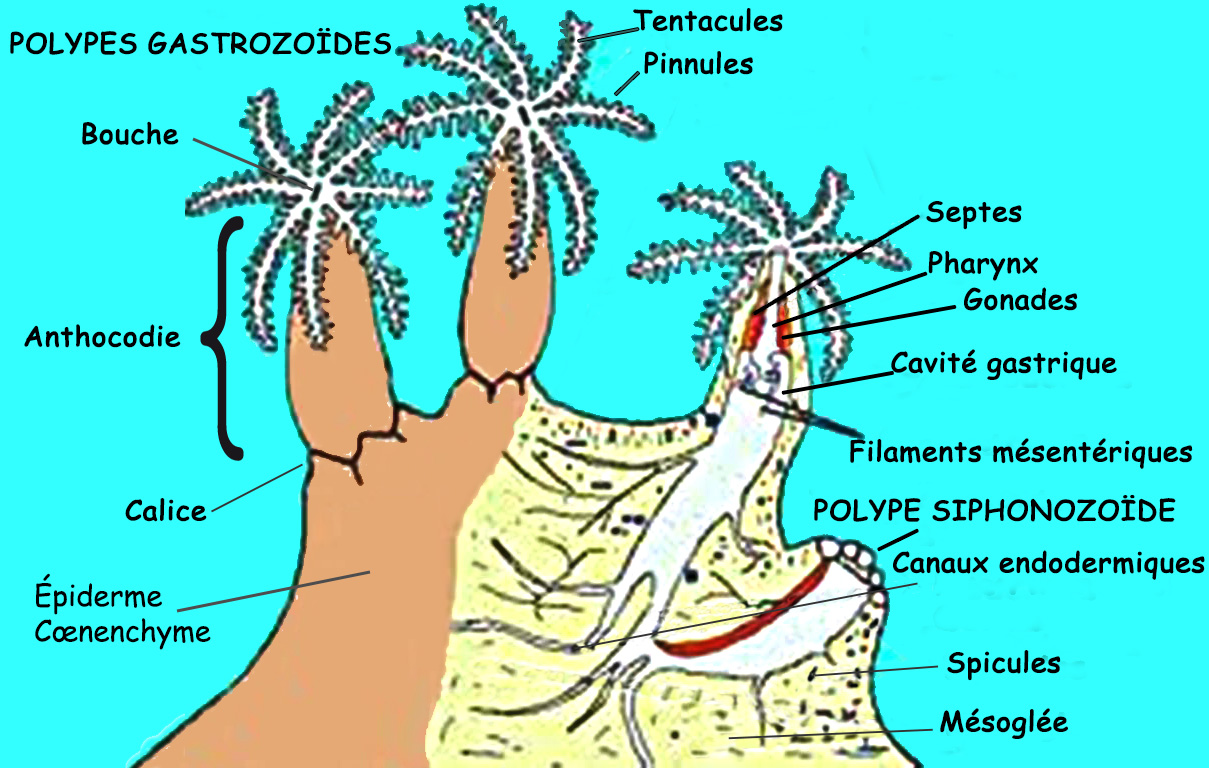
2.3.1. Polypes autozoïdes
Tous les Octocoralliaires disposent de polypes autozoïdes (ou gastrozoïdes). Dédiés essentiellement au nourrissage ils assurent les fonctions de capture, de digestion des aliments, de respiration, de captation de lumière (tel un puit de lumière) vers le cœur de la colonie ainsi que la reproduction puisqu’ils portent les gonades (ovaires et testicules). La capture prend ici un sens étendu, assurée de plusieurs manières, comme on le verra plus tard.
2.3.1.1. Anthocodie, partie émergeante

La partie émergeante du polype (l’anthocodie), de forme tubulaire plus ou moins allongée, de quelques millimètres à plus de dix centimètre de longueur chez certaines espèces comme Xenia, est prolongée de ses huit tentacules. Entre les tentacules, se situe la bouche ou débute le pharynx. A la base de l’anthocodie, les stolonifères développent une partie en saillie formant un calice ou anthostèle, plus épaisse, dure et chargée de spicules. Contrairement à une idée répandue, l’anthostèle ne se limite pas à la partie émergeante, elle inclut tout le canal gastrodermique qui s’enfonce parfois profondément dans le corps. Selon les espèces, la partie supérieure de l’anthocodie, molle, peut se contracter dans le calice par simple déflation ; on dit que le polype est contractile. L’anthocodie peut également s’invaginer entièrement ne laissant apparaître qu’un orifice, on dit alors qu'il est rétractile. Chez certaines espèces comme Anthelia spp., le polype ne peut pas se contracter ni se rétracter. Chez d'autres, les polypes sont seulement formés d’une longue anthocodie, simplement déprimée à la base, juste au-dessus du stolon. Les Pennatules s’égarent de ce schéma habituel : elles disposent d’un polype principal (le rachis) d’où partent des rangées de polypes secondaires. Quelques espèces comme Sarcophyton spp., disposent de longs polypes en mesure de se rétracter profondément à l’intérieur du corps. Ces derniers permettraient de conduire la lumière vers les couches internes pourvues de zooxanthelles.
2.3.1.2. Tentacules et pinnules
Sauf exception, les tentacules sont au nombre de huit. De 1 à 50 mm, ils se répartissent autour du disque oral de chaque polype et alternent avec les huit cloisons. De forme parfois asymétrique, comme chez les pennatules, ils sont creux, effilés à leur extrémité et en général, portent des petites ramifications latérales, les pinnules. Contractiles, ils peuvent être épanouis ou rétractés, le jour ou la nuit, selon l’espèce. Les tentacules, mobiles, tactiles et préhensiles, contiennent de nombreux cnidocytes et interviennent dans la défense, la capture des proies et la nutrition. Ils assurent également l'acheminement de la nourriture vers la bouche.
2.3.1.3. Disque oral et bouche
La bouche, de forme ovale au centre du disque oral, présente à l’une de ses extrémités étroites un élargissement indiquant la position du siphonoglyphe (voir ci-après) qui se prolonge à l’intérieur du pharynx.
2.3.1.4. Pharynx
Le pharynx ou stomodeum, ovale, peut être lisse ou parcouru de sillons. Il s’étend plus bas jusqu’à la cavité gastrique. Il est parcouru à son extrémité ventrale par une gouttière bordée de cellules ciliées, le siphonoglyphe (absent chez les scléractiniaires). Cette gouttière est destinée à assurer la circulation de l’eau vers l'intérieur, dans la cavité gastrique et participerait à l’ingestion de particules alimentaires.
2.3.1.5. Cavités gastriques

La cavité gastrovasculaire (ou cœlenteron) de chaque polype est formée d'un ensemble de huit cavités gastriques, indépendantes les unes des autres. Dans la partie supérieure, elles sont soudées entre elle par des cloisons ou septes, qui se poursuivent longitudinalement, délimitant ainsi depuis le disque oral et au niveau du pharynx, huit compartiments distincts (les chambres radiaires), communiquant directement avec les sillons tentaculaires. Il y a une chambre asulcale (sans sillon), une chambre sulcale (avec sillon) et six chambres latérales. Au-dessous du pharynx, les cavités se prolongent vers la base de la colonie sous forme de canaux longitudinaux endodermiques terminés en cul de sac. A ce niveau, les bords externes des cloisons (cloisons mésentériques) sont reliés à la paroi des polypes, alors que les bords internes sont libres et portent des cordons épais et onduleux, les filaments mésentériques.
On distingue huit filaments mésentériques, dissemblables tant par leur constitution que par leur origine : les deux filaments asulcaux sont longs, rectilignes souvent bilobés et descendent jusqu’au fond du polype. Fortement ciliés, ils servent à créer un courant d’eau exhalant, vers la bouche. Les six autres filaments (deux sulcaux et quatre latéraux) sont plus réduits, ils contiennent des glandes digestives et achèvent la digestion des particules alimentaires commencée dans le pharynx. En général ces derniers produisent les gonades.
Dans les colonies massives, une quantité parfois importante de polypes se prolonge profondément dans la masse en un tube nommé canal gastrodermique. Ce canal est également connecté aux autres parties de la colonie par un réseau communicant le solenia. Dans les colonies qui disposent de tissus fins autour d’un axe ou celles qui s’étendent sur le substrat, la cavité gastrique est réduite.
2.3.1.6. Circulation d’eau
La circulation de l’eau au sein des colonies coralliennes, joue un rôle fondamental dans le métabolisme du corail : elle facilite la nutrition, permet les échanges gazeux et favorise l'évacuation des déchets du métabolisme.
Il existe au niveau de la bouche des polypes gastrozoïdes et des polypes siphonozoïdes, deux courants ciliaires plus ou moins puissants :
d
’un côté, un courant inhalant provient du siphonoglyphe et de l’autre, les filaments mésentériques asulcaux génèrent un courant ciliaire exhalant ascendant, depuis la base jusqu’au sommet de la cavité gastrovasculaire. Leurs actions conjuguées assurent l’arrivée d'une grande quantité d’eau dans les polypes et son expulsion, tout en permettant aux particules nutritives de pénétrer et éventuellement, de ressortir de l’orifice buccal.
2.3.2. Polypes siphonozoïdes

Les espèces aux formes massives comme celles de la famille Pennatulidae, quelques alcyoniidés (Sarcophyton, Lobophyton, Hetroxenia…) et les gorgonidés sont dimorphes : elles possèdent en plus des polypes autozoïdes, un autre type de polypes dits siphonozoïdes.
Ces derniers, démunis de tentacule et dont on ne voit que les orifices à fleur de l'épiderme, ne sont pas rétractiles. Ils sont spécialisés dans les échanges respiratoires de la colonie.
Leurs cloisons gastriques et leurs filaments mésentériques son plutôt atrophiées, alors que les siphonoglyphes sont très développés.
Les filaments mésentériques asulcaux (sans sillon) assurent la circulation d’eau, l'exhalant de bas en haut. Le courant inhalant qui en résulte assure la respiration jusqu'au sein du polype ainsi que le maintien d'une pression interne permettant aux colonies massives ou peu arborescentes, de se dresser. Certaines espèces ne forment d’ailleurs des polypes siphonozoïdes qu’au-delà d’une certaine taille.
2.4. Squelette et hydrosquelette

La grande majorité des Octocoralliaires possède un squelette interne (endosquelette) composé d'éléments en carbonate de calcium (calcite) et organique, de petites tailles : les sclérites (terme employé au masculin ou au féminin) ou spicules (masculin), les scientifiques n'ayant pas tranché sur le terme le plus approprié, répartis à l’intérieur du cœnenchyme. Ces sclérites plus ou moins acérés assurent le maintien du corail et contribuent à sa protection contre des prédateurs. L’aspect des sclérites est très varié. En forme d’aiguille plus ou moins longue de 0.02 à 4 mm, plus ou moins épaisse, rectiligne ou au contraire difforme, en forme de boutons épineux, de croix, d’haltères, d’étoiles ou de massues.
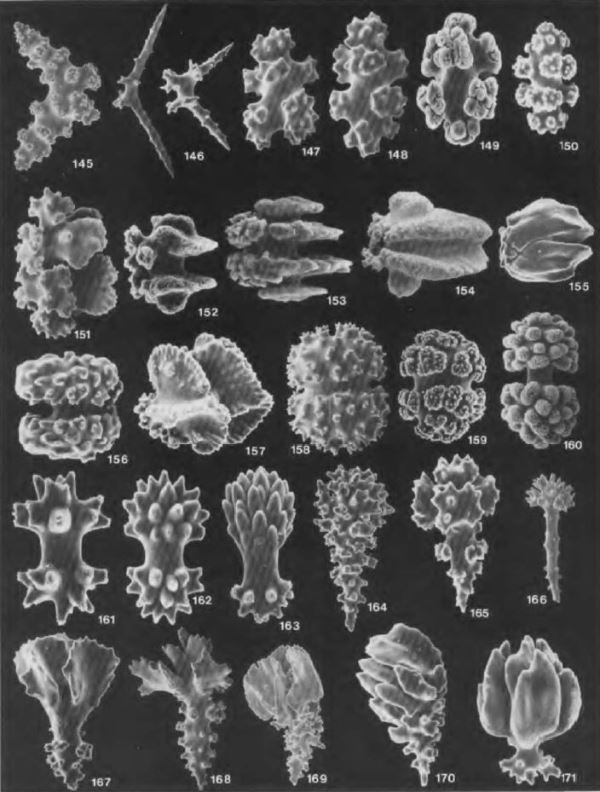
Il est bien souvent impossible de déterminer visuellement un Octocoralliaire au-delà du genre. Ce n’est pas vraiment un handicap pour la pratique du hobby récifal. Les scientifiques poussent l’identification jusqu’à l’espèce par une analyse microscopique des sclérites, de leur disposition ou de leur forme. Des catalogues (photo ci-contre) répertorient les formes aux noms évocateurs : papillon, rayon, disque, baril double tête, double étoile, torche, feuille... Leur forme variant selon leur localisation sur l’animal, les prélèvements sont effectués à la fois à la base des colonies et sur les ramifications. L’arrangement des sclérites de la colonie est également un indicateur pour leur identification. Certaines espèces de Nepthea sont empaquetées dans un réseau de larges sclérites. Des sclérites en forme d’arcs allongés peuvent être organisées en collerettes ou couronnes renforçant le calice ou en pointes à la base des tentacules (Illustration 2)
Pour analyser un Octocoralliaire il faut, dans un premier temps, découper au scalpel un échantillon du tissu du corail, dissoudre la matière organique avec quelques gouttes d’hypochlorite de sodium (eau de javel), laver plusieurs fois les sclérites à l’eau distillée, les laisser sécher sur une lame de microscope puis les recouvrir d’une lamelle. Il est parfois nécessaire de les consolider dans une inclusion de résine époxy. Les spicules mises en évidence, on peut les observer sous grossissement moyen. Mais à ce stade, l'identification nécessite de la persévérance : contrairement aux scléractiniaires, les rapports de recherches sont peu accessibles, les différences entre espèces sont parfois ténues, d'autant plus que celles vivant en captivité peuvent développer des malformations. Mais pas de panique en cas d’erreur : la classification est en plein remaniement et promet quelques bouleversements dans les prochaines décennies.

Chez certains groupes (pennatules, gorgones), le squelette prend la forme d’un axe semi-rigide, formé de matière protéinique et de substances collagènes, à l’aspect de la corne (la gorgonine), enrobé d’un fin tissu, le cœnosarque appelé cortex chez les gorgones (ex. gorgones des familles Holaxonia et Calcaxonia). Les plus complexes incorporent dans la composition de leur axe (la médulla), des matières calcaires plus ou moins agrégées : cimentées comme chez le fameux Corail rouge de Méditerranée Corallium rubrum ou libres comme chez les gorgones du sous-ordre Scleraxonia qui incluent des sclérites indépendantes. Ces structures plus ou moins souples possèdent des médullas fortement liées, compactes, solides et peuvent parfois dépasser les trois mètres d’envergure. D’autres moins complexes, constituées de spicules non agglomérées dans la gorgonine, sont bien plus fragiles.
Les espèces de la famille Briareidae, qui se développent en forme de membranes encroûtantes, sont considérées comme des gorgones dont la médulla cornée a migré et s‘est étendue sur la face d’appui, le cortex accueillant les polypes étant sur la face supérieure.
Chez quelques stolonifères comme Tubipora, le squelette prend la forme d’une enveloppe cornée dans laquelle le polype peut se rétracter. Certaines espèces du genre Sinularia peuvent agglomérer des sclérites à leur base, pour former une structure rigide, rocheuse, la spiculite.
De nombreux Octocoralliaires n’ont pas de squelette, leur maintien est alors assuré par un réseau de canaux sous pression d’eau. Cet hydrosquelette peut se dégonfler quasi instantanément, en réponse à une perturbation ou à un stress. Le corail se vide alors de son eau par la bouche pour rapetisser en moins d’une minute. Les Octocoralliaires contenant peu ou pas de squelette sont en général de petites tailles, mais on trouve des colonies supérieures à un mètre dont le corps contient de véritables sacs à eau.

2.5. Musculature et système nerveux
Les septes abritent de puissants muscles longitudinaux endodermiques permettant la rétraction de la partie supérieure du polype, et d’autres, transversaux et moins développés, servant à mouvoir les tentacules. Ces muscles sont commandés par un système nerveux.
La mésoglée contient un réseau de fibres nerveuses très primitif (deux plexus nerveux, un sous-ectodermique et un sous-endodermique). Les quelques centaines de milliers de neurones, également dispersées dans l’endoderme et l’ectoderme de manière à détecter les proies de toutes parts, sont plus denses autour de la bouche. Ces cellules nerveuses répondent aux stimulations et transmettent des impulsions aux cellules contractiles.
2.6. Substances bioactives
Bien que les cnidaires soient des organismes relativement simples, leur métabolisme produit des substances bioactives complexes que l'on découvre encore. Ces substances, appelées métabolites secondaires, sont tout sauf des sous-produits. Leur production nécessite des enzymes spécifiques exigeant beaucoup d'énergie de leur part. Destinées à dissuader des prédateurs, à nettoyer les envahisseurs, à lutter pour la conquête de l'espace, à se protéger des radiations solaires ou combattre les maladies, elles jouent un rôle physiologique et écologique si important pour la survie de l’animal, et donc sa maintenance en captivité, que nous ne pouvons les passer sous silence. Nous aborderons plus en détail cet aspect de l'écologie chimique des Octocoralliaires, dans un chapitre ultérieur.
Références
- K. Fabricius, P. Alderslade. Soft corals and sea fans – A comprehensive guide to the tropical shallow-water genera of the Central-West Pacific, the Indian Ocean and the Red Sea. Australian Institute of Marine Science, Townsville, Australia. 2001. 264 p
- J. Sprung, J. C. Delbeck. L’aquarium récifal, volume 2. Ricordea Publishing 1999.
- J. Vimal. Physiopathologie des Coraux. Thèse Université Claude-Bernard. Lyon I, mai 2007.
- F. M. Bayer, M. Grasshoff, Jakob Verseveldt, Illustrated Trilingual Glossary of Morphological and Anatomical Terms Applied to Octocorallia, E. J. BRILL , DR. W. BACKHUYS LEIDEN, The Netherland, 1983.
- Les Octocoralliaires – Mer et littoral
- M. P. Janes. Laboratory methods for the identification of soft corals (Octocorallia: Alcyonacea). Advances in Coral Husbandry in Public Aquariums. Public Aquarium Husbandry Series, vol. 2. R.J. Leewis and M. Janse (eds.),ch 46, pp. 413-426. 2008 Burgers’ Zoo, Arnhem, the Netherlands.
Tous mes remerciements aux auteurs des photos pour leur accord de publication et à mes correcteurs Christian SEITZ, Olivier SOULAT et Jérémie VIDAL DUPIOL, pour leur soutien.
Denis TOURNASSAT
Article publié par Cap récifal le 1 novembre 2014 avec l'aimable autorisation de l'auteur.
Modifié par Denisio
Commentaires recommandés
Il n’y a aucun commentaire à afficher.